



 Рейтинг: 4.4/5.0 (1839 проголосовавших)
Рейтинг: 4.4/5.0 (1839 проголосовавших)Категория: Бланки/Образцы
Эколого-флористическая классификация основана на дедуктивно-индуктивном подходе, при котором главную роль играет процесс установления синтаксонов «снизу». Это происходит путем группирования сообществ по сходству флористического состава, который в значительной степени отражает экологические условия и стадию сукцессии. При этом упор делается на том, что различные виды по-разному связаны с различными условиями произрастания, то есть с различными фитоценозами, формирующимися в данных условиях. Свойство видов быть более или менее прочно связанными с определенным типом фитоценозов Браун-Бланке называет верностью. Им различается пять уровней верности:
Верные виды (степень верности 5) – виды, исключительно или почти исключительно связанные с определенным типом сообществ.
Прочные виды (степень верности 4) – виды, встречающиеся и в других сообществах, но более редко и в меньшем числе или с пониженной жизненностью.
Благосклонные виды (степень верности 3) – виды, которые встречаются более или менее обильно во многих фитоценозах, но оптимальное их развитие связано с одним определенным типом сообщества.
Неопределенные виды (степень верности 2) – виды без явно выраженной связи с каким-либо определенным типом сообществ.
Чуждые виды (степень верности 1) – редкие и большей частью случайные включения из других растительных сообществ или же реликты ранее бывших растительных сообществ.
Система Браун-Бланке предполагает, что каждое растительное сообщество экологически специфично и данная специфичность проявляется во флористическом наборе, уникальном для каждого конкретного типа местообитания (рис. 1). Виды, характерные для конкретного синтаксона, называются диагностическими. Они, в зависимости от уровня верности разделяются на три группы:
Характерные виды встречаются только в одном синтаксоне или встречаются в этом синтаксоне чаще, чем в других благодаря своей экологической специализации. Характерные виды могут быть выявлены только при наличии большого количества описаний, охватывающих основные типы фитоценозов в пределах достаточно большого района. К характерным видам при этом относятся верные, прочные и благосклонные виды. Как правило, это стенотопные виды.
Дифференцирующие виды диагностируют синтаксон границей своего ареала и входят в состав нескольких синтаксонов.
Константные виды встречаются с высоким постоянством и часто с высоким обилием, но проходят через этот синтаксон «транзитом». Виды, константные для низших единиц классификации, могут быть дифференцирующими или характерными для более высоких единиц классификации.

Рис. 1. Схема, показывающая основные варианты диагностических видов в системе Браун-Бланке:
А, Б, В – ассоциации; а, б, в – субассоциации; 1, 2 – характерные виды ассоциации Б; 3, 4 – характерные виды ассоциации А; 5 – характерный вид ассоциации В; 4, 6 – дифференциальные виды субассоциации а; 7, 8 – дифференциальные виды субассоциации в; субассоциация б выделяется по критерию отсутствия дифференциальных видов субассоциаций а и в; 9, 10, 11 – константные виды, которые могут быть характерными для классификационных единиц более высокого ранга (Миркин и др. 2002)
Основной таксономической категорией, или основной единицей растительности, в системе Браун-Бланке является ассоциация (табл. 1). Ассоциация школы Браун-Бланке – единица крупная, широкого объема, подразделяемая в свою очередь на более мелкие единицы, в то время как при доминантной классификации это единица узкого объема, зачастую являющаяся низшей единицей системы классификации. Она выделяется на основании анализа всего флористического состава и отличается свойственными ей характерными видами, которые могут быть верными, прочными или же только благосклонными.
Вышестоящие таксоны, относящиеся к союзам, порядкам, классам, имеют свои характерные виды. Чем выше ранг, тем большим числом характерных видов обладает данный таксон. Класс – это единица с выраженной собственной физиономией, флористическая комбинация при установлении класса стоит на втором месте. Порядок – крупный вариант класса, устанавливаемый на основе флористических критериев. Союз чаще всего – географический или экологический вариант порядка.
Таблица 1. Обзор фитоценотических единиц системы Браун-Бланке.






















Выпуск № 1-3 / том 12 / 2010
КодыПроведена оптимизация методики оценки сырьевой продуктивности лекарственных растений по их проективному покрытию в растительных сообществах, описанных в системе единиц классификации Браун-Бланке. Метод использован для оценки продуктивности лекарственных видов в растительных сообществах Южного Урала.
Abstract 2010 year, VAK speciality — 03.00.00, author — Fedorov N. I. Zhigunova S. N. Mihaylenko O. I. Samoylova L. YU.Optimization of methodology of estimation the raw efficiency of drug plants on their projective covering in the vegetative communities described in system of Braun-Blanquet classification is lead. The method is used for estimation the efficiency of medicinal kinds in vegetative communities of Sonth Urals.
Научная статья по специальности " Биология " из научного журнала "Известия Самарского научного центра Российской академии наук", Федоров Н. И. Жигунова С. Н. Михайленко О. И. Самойлова Л. Ю.Скопируйте отформатированную библиографическую ссылку через буфер обмена или перейдите по одной из ссылок для импорта в Менеджер библиографий.
Федоров Н. И. Жигунова С. Н. Михайленко О. И. Самойлова Л. Ю. Методика оценки продуктивности лекарственных видов в растительных сообществах, описанных в системе единиц эколого-флористической классификации Браун-Бланке // Известия Самарского научного центра РАН. 2010. №1-3. URL: http://cyberleninka.ru/article/n/metodika-otsenki-produktivnosti-lekarstvennyh-vidov-v-rastitelnyh-soobschestvah-opisannyh-v-sisteme-edinits-ekologo-floristicheskoy (дата обращения: 01.10.2016).
Федоров Н. И. et al. "Методика оценки продуктивности лекарственных видов в растительных сообществах, описанных в системе единиц эколого-флористической классификации Браун-Бланке" Известия Самарского научного центра Российской академии наук 12 (2010). URL: http://cyberleninka.ru/article/n/metodika-otsenki-produktivnosti-lekarstvennyh-vidov-v-rastitelnyh-soobschestvah-opisannyh-v-sisteme-edinits-ekologo-floristicheskoy (дата обращения: 01.10.2016).
Федоров Н. И. Жигунова С. Н. Михайленко О. И. & Самойлова Л. Ю. (2010). Методика оценки продуктивности лекарственных видов в растительных сообществах, описанных в системе единиц эколого-флористической классификации Браун-Бланке. Известия Самарского научного центра Российской академии наук URL: http://cyberleninka.ru/article/n/metodika-otsenki-produktivnosti-lekarstvennyh-vidov-v-rastitelnyh-soobschestvah-opisannyh-v-sisteme-edinits-ekologo-floristicheskoy (дата обращения: 01.10.2016).
Скопируйте отформатированную библиографическую ссылку через буфер обмена или перейдите по одной из ссылок для импорта в Менеджер библиографий.
Федоров Н. И. Жигунова С. Н. Михайленко О. И. Самойлова Л. Ю. Методика оценки продуктивности лекарственных видов в растительных сообществах, описанных в системе единиц эколого-флористической классификации Браун-Бланке // Известия Самарского научного центра РАН. 2010. №1-3 С.846-849.
Федоров Н. И. et al. "Методика оценки продуктивности лекарственных видов в растительных сообществах, описанных в системе единиц эколого-флористической классификации Браун-Бланке" Известия Самарского научного центра Российской академии наук 12 (2010).
Федоров Н. И. Жигунова С. Н. Михайленко О. И. & Самойлова Л. Ю. (2010). Методика оценки продуктивности лекарственных видов в растительных сообществах, описанных в системе единиц эколого-флористической классификации Браун-Бланке. Известия Самарского научного центра Российской академии наук
Школьники описывают типичный участок луговой растительности - обычная геоботаническая практика. В данном случае нет необходимости закладывать пробную площадку. Фото с сайта www.ecosystema.ru/
Разбирается история и принципы одного из методов геоботанического описания - метод Браун-Бланке. Это метод классификации растительного покрова, предполагающий выделение типичных растительных комплексов по большому набору совместно произрастающих видов растений. Описание таких растительных ассоциаций предполагает дальнейший анализ их связи с различными абиотическими и биотическими условиями их произрастания. Хотя выделение однородных ассоциаций - это в большой мере субъективный и условный процесс, что снижает его фальсифицируемость, но авторы статьи видят за ним будущее развитие геоботаники в России. Актуальность и перспективность ему придают массовость ботанических определений того или иного района, независмость классификации от сопутствующих абиотических факторов.
Читателям Журнала Общей Биологии предлагается краткий обзор истории развития одного из методов классификации растительности, применяемый в фитоценологии. Этот метод называется по имени своего создателя, швейцарца Жозья Браун-Бланке, изучавшего в начале 20-го века растительность Европы. Обзор написан Б.М.Миркиным из Института Биологии УНЦ РАН и Л.Г.Наумовой из Башкирского государственного педагогического Университета г.Уфы.
Метод Браун-Бланке - это способ классификации растительных ассоциаций. В его основе лежит экспертное выделение более или менее единообразных комплексов растений. Каждый комплекс характеризуется, как правило, диагностическими видами, и их может быть довольно много. При описании кмплекса ему дается название, которое, как и в таксономии, состоит из двух наменований. Описание сопровождается указанием различных характеристик почв, фаунистических элементов и т.д. Авторы статьи указывают, что в России метод Браун-Бланке начал применяться довольно поздно, только в 70-х годах 20-го века. Это было связано с тем, что россиийская геоботаническая школа развивала т.н. метод доминантов. В статье он назван 'еловая догма'. Несмотря на некоторый уничижительный оттенок, придаваемый словом 'догма', эта метафора довольно точно передает направленность российских геоботанических разработок 20-го века. Основные работы в СССР были сделаны на базе изучения разнообразных лесных массивов. А для лесов вид-эдификатор или вид-доминант является, наряду с абиотическими характеристиками, ведущим фактором, формирующим облик растительности. Однако при расширении объектов исследования до луговой и степной растительности метод доминантов перестает работать, хотя бы из-за того, что выделение доминирующих видов превращается в довольно трудную задачу. Ведь по сравнению с лесной растительностью распределение видовых богатств становится более равномерным. К сообществам со смещенным доминированием относятся и синантропные фитоценозы. Влияние антропогенного фактора и этапы восстановления подобных сообществ были более или менее удачно показаны с использованием именно метода Браун-Бланке (см. статью в ЖОБ. 2007 'Синантропные растительные сообщества: модели организации и особенности классификации '); обычный метод доминантов к нарушенным сообществам не применим. В силу всех этих причин в России постепенно стал входить в научный обиход метод Браун-Бланке.
Его применение активизировалось в 90-х годах 20-го века, так как именно в это время начали создаваться глобальные базы данных, в том числе и ботанические. Базы данных выполняли задачу каталогизации растительности в разных странах и на планете в целом. Описание растительных ассоциаций по методу Браун-Бланке как нельзя лучше подходило для составления таких реестров, так как по определению включало большое число видов растений.
Миркин и Наумова обращают внимание на важную деталь фито-ценологической классификации растительности. Изначально, при разработке метода, полагалось считать, что эксперт выделяет целостную и дискретную группировку растительности. Именно такой дискретной группировке и полагалось присваивать наименование в рамках синтаксономии (то самое двойное название). Сами авторы этой статьи являются континуалистами (см. статьи в ЖОБ: 2004 'Значение классификации растительности для современной экологии ', 2006, 'Континуальность и дискретность живого покрова: проблема масштаба '), поэтому выделение устойчивых ассоциаций становится в большой степени условным. Оно превращается в удобный (для конкретного исследователя!) описательный инстумент. Экологическую значимость этим выделенным 'удобным' единицам придает дальнейший анализ взаимосвязи ассоциаций с теми или иными параметрами внешней среды.
Использование этого инструмента обосновано пятью причинами: его необходимостью, доступностью, информативностью, единообразием, оптимальностью. Необходимость подразумевает обязательность расчленения целого на составные части при анализе целого. Доступность - это возможность оценки флористического облика без специальных инструментов, достаточно знать таксономию растений. Под информативностью авторы понимают возможность определить стадию сукцессии и изменение условий среды по облику растительности; отметим, что информативность в данном случае является результатом дальнейшего экологического анализа, который может быть проделан, а может быть опущен, так как метод Браун-Бланке - всего лишь способ классификации растительности. Единообразие обеспечивается коллективным соглашением, выработанным на основе интуитивного понимания учеными, какую ассоциацию считать однородной. Наконец, оптимальность понимается как выбор типичной геоботанической площадки в противовес случайной площадке (или набору случайных площадок) при классическом геоботаническом описании. Это позволяет сэкономить силы исследователей, не обращать внимания на эпизодические и случайные нарушения растительного покрова местообитаний.
Б.М.Миркин и Л.Г.Наумова разбирают критику метода, высказанную А.М.Гиляровым, экологом из МГУ, биофак. Гиляров считает, что классификация по Браун-Бланке не имеет экологического смысла, и потому является рудиментом естественной истории. Подобные рудименты сильно замедляют развитие современной экологии. В сущности возражения Гилярова сводятся к неопределенности правил и критериев классификации, изначально не несущих никакого экологического смысла. К негативным сторонам метода он относит также условность и субъективность выделения однородных единиц, нечеткость меры однородности растительных ассоциаций. Ответ авторов статьи сводится к следующему. Данная классификация всего лишь описывает портрет растительного покрова без привлечения дополнительных экологических критериев. Если привлечь любые абиотические или экологические критерии, то мы уже заранее заложим определенную интерпретацию.
С условностью и субективностью выделения растительных синтаксонов авторы согласны. Это порождает большую путаницу, беспрестанную ревизию синтаксонов и необходимость указывать не только автора синтаксона, но и год его установления, чтобы понятен был смысл и объем синтаксона. Однако субъективность синтаксона - это естественное следствие континуальности растительного покрова и, соответственно, множественности возможных решений.
Еще по темам: Геоботаника. МетодологияA. С. Желтухин, Ю. Г. Пузаченко, И. П. Котлов, И. И. Широня, Р. Б. Сандлерский
Пространственно-временная динамика следовой активности лесной куницы (Martes martes L.), зайца-беляка (Lepus timidus L.) и белки (Sciurus vulgaris L.) в европейской южной тайге
Стр. 262–283
Резюме. Abstract
Синопсис: Ежедневники лесных обитателей можно прочитать зимой
Н. Б. Ермаков, А. В. Ларионов, М. А. Полякова, Ю. В. Плугатарь
Экологическая интерпретация высших единиц степной растительности в горах юга Средней Сибири методом количественной ординации
Стр. 293–302
Резюме. Abstract
Синопсис: Три пишем, два в уме: единицы геоботанического описания и экологические параметры
B. Б. Мартыненко, П. С. Широких, Б. М. Миркин, Л. Г. Наумова, Э. З. Баишева, А. А. Мулдашев
Синтаксономический анализ влияния инициальной стадии на вторичную автогенную сукцессию широколиственного леса
Стр. 303–313
Резюме. Abstract
Д. О. Логофет, И. Н. Белова, Е. С. Казанцева, В. Г. Онипченко
Ценопопуляция незабудочника кавказского (Eritrichium caucasicum ) как объект математического моделирования. I. Граф жизненного цикла и неавтономная матричная модель
Стр. 106–121
Резюме. Abstract
А. А. Шулаков, А. В. Егоров, В. Г. Онипченко
Филогенетический анализ растительных сообществ высокогорий Тебердинского заповедника
Стр. 135–144
Резюме. Abstract
Синопсис: В каком случае родственники - лучшие соседи?
В. В. Акатов, Т. В. Акатова, Т. Г. Ескина, Ю. С. Загурная, Н. М. Сазонец, С. Г. Чефранов
Эффект компенсации плотностью в маловидовых растительных сообществах материковых территорий (на примере Западного Кавказа)
Стр. 38–53
Резюме. Abstract
М. Л. Бутовская, Е. В. Веселовская, К. В. Левина, В. В. Ростовцева
Механизмы репродуктивного поведения человека: визуальные маркеры мужской привлекательности, их связь с ольфакторными маркерами, сексуальным опытом и фазой месячного цикла у женщин-экспертов
Стр. 63–77
Резюме. Abstract
Синопсис: Научный глянец: как выбрать своего мужчину
Д. О. Логофет, Н. Г. Уланова, И. Н. Белова
Поливариантный онтогенез у вейников: новые модели и новые открытия
Стр. 438–460
Резюме. Abstract
А. М. Кипкеев, О. В. Чередниченко, Д. К. Текеев, В. Г. Онипченко
Скорость микросукцессий: восстановление структуры и флористической насыщенности альпийских фитоценозов после реципрокных пересадок участков дернины
Стр. 461–474
Резюме. Abstract
Синопсис: Восстановление структуры альпийских фитоценозов после пересадок участков дернины
Б. М. Миркин, Л. Г. Наумова
Вклад теоретического наследия Л.Г. Раменского в современную науку о растительности (к 130-летию со дня рождения ученого)
Стр. 244–256
Резюме. Abstract
Д. О. Логофет, О. И. Евстигнеев, А. А. Алейников, А. О. Морозова
Сукцессия, вызванная жизнедеятельностью бобра (Саstor fiber L.): II. Уточненная марковская модель
Стр. 126–145
Резюме. Abstract
Б. М. Миркин, Л. Г. Наумова
Концепция растительного сообщества: история и современное состояние
Стр. 63–76
Резюме. Abstract
О. Ю. Конева
В популяции моллюсков Lymnaea stagnalis из радиационно-неблагополучною региона обнаружено двукратное увеличение содержания ДНК в гемоцитах
Стр. 466–477
Резюме. Abstract
Синопсис: «Кометы» и улитки могут указывать на повышение радиационной нагрузки
В. Б. Мартыненко, П. С. Широких, Б. М. Миркин, Л. Г. Наумова
Синтаксономический анализ восстановительных сукцессии после вырубки светлохвойных лесов Южно-Уральского региона
Стр. 478–490
Резюме. Abstract
Л. Э. Ялковская, А. В. Бородин, М. А. Фоминых
Модульный подход к изучению флуктуирующей асимметрии комплексных морфологических структур у грызунов на примере нижней челюсти рыжей полевки (Clethrionomys glareolus. Arvicolinae, Rodentia)
Стр. 385–393
Резюме. Abstract
Синопсис: Одна кость – две судьбы
A. В. Почикалов, Д. В. Карелин
Окологодичные наблюдения за разложением опада тундровых растений через потерю массы и эмиссию СО2: роль биотических и абиотических факторов, сезонов года, биотопа и пространственно-временного масштаба
Стр. 163–181
Резюме. Abstract
Синопсис: Диалектика распада или как масштаб меняет выводы
Э. Д. Владимирова, В. В. Морозов
Формальное описание поведения млекопитающих по материалам зимних троплений на примере лесной куницы (Martes martes)
Стр. 182–203
Резюме. Abstract
Б. М. Миркин, Л .Г. Наумова
Основные тенденции развития теории синтаксономии и динамики растительных сообществ
Стр. 83–94
Резюме. Abstract
Синопсис: Геоботаника ждет своего Хеннига
В. Г. Суховольский, Т. М. Овчинникова, С. Д. Бабой
Высотно-поясная зональность древесных пород в горных условиях Саян: модель экологических фазовых переходов второго рода
Стр. 38–47
Резюме. Abstract
Р. К. Аджиев, В. Г. Онипченко, Д. К. Текеев
Сохранение жизнеспособности погребенных семян в альпийских фитоценозах северо-западного Кавказа: итоги пятилетнего эксперимента
Стр. 453–458
Резюме. Abstract
Л. В. Недорезов
Анализ экспериментальных временных рядов Г. Ф. Гаузе с помощью моделей с непрерывным временем
Стр. 114–124
Резюме. Abstract
Н. Ю. Феоктистова, О. Ф. Чернова, И. Г. Мещерский
Декоративные формы хомячков рода Phodopus (Mammalia, Cricetinae) - анализ распространения генетических линий и особенности изменения волосяного покрова
Стр. 138–154
Резюме. Abstract
Синопсис: Зачем нам знать, откуда взялись домашние хомячки?
A. А. Поздняков
Структура морфологической изменчивости (на примере морфотипов жевательной поверхности первого нижнего коренного зуба серых полевок)
Стр. 127–139
Резюме. Abstract
Синопсис: Гомологическая изменчивость у полевок – результат эпигенетической регуляции.
Д.Б. Гелашвили, Е.В. Чупрунов, М.О. Марычев, Н.В. Сомов, А.И. Широков, А.А. Нижегородцев
Приложение теории групп к описанию псевдосимметрии биологических объектов
Стр. 497–513
Резюме. Abstract
Синопсис: Посчитанные отражения
Д.Б. Гелашвили, В.И. Якимов, Д.И. Иудин, Г.С. Розенберг, Л.А. Солнцев, С.В. Саксонов, М.С. Снегирева
Фрактальные аспекты таксономического разнообразия
Стр. 115–130
Резюме. Abstract
Б.М. Миркин, В.Б. Мартыненко, П.С. Широких, Л.Г. Наумова
Анализ факторов, определяющих видовое богатство сообществ лесов Южного Урала
Стр. 131–143
Резюме. Abstract
Синопсис: Видовое богатство лесов Южного Урала
И.Я. Павлинов
Замечания о биоморфике (экоморфологической систематике)
Стр. 187–192
Резюме. Abstract
B. А. Лухтанов, В. Г. Кузнецова
Молекулярно-генетические и цитогенетические подходы к проблемам видовой диагностики, систематики и филогенетики
Стр. 415–437
Резюме. Abstract
Синопсис: Возможности и ограничения современных методов филогенетики
Б. М. Миркин, С. М. Ямалов, А. В. Баянов, Л. Г. Наумова
Вклад метода Браун-Бланке в объяснение причин видового богатства растительных сообществ
Стр. 285–295
Резюме. Abstract
Синопсис: Антропогенный фактор увеличивает видовое разнообразие
В. В. Акатов, Т. В. Акатова, С. Г. Чефранов, А. Е. Шадже
Уровень полночленности и потенциал инвазибельности растительных сообществ: гипотеза соотношения видовых фондов
Стр. 328–340
Резюме. Abstract
Синопсис: Гостеприимство фитоценозов
А. Г. Васильев
Феногенетическая изменчивость и популяционная мерономия
Стр. 195–209
Резюме. Abstract
И. Я. Павлинов, О. Г. Нанова
К изучению морфологического разнообразия размерных признаков черепа млекопитающих. 3. Дистантный анализ объема и заполнения морфопространства
Стр. 35–55
Резюме. Abstract
Синопсис: Как измерить морфологическое пространство?
А. А. Лисовский, И. Я. Павлинов
К изучению морфологического разнообразия размерных признаков черепа млекопитающих. 2. Скалярные и векторные характеристики форм групповой изменчивости
Стр. 428–433
Резюме. Abstract
Синопсис: Новые математические методы изучения групповой изменчивости
Н. Г. Уланова, И. Н. Белова, Д. О. Логофет
О конкуренции среди популяций с дискретной структурой: матричная модель динамики популяций вейника и березы, растущих совместно
Стр. 441–457
Резюме. Abstract
Синопсис: Когда березы были еще маленькие.
И. Я. Павлинов. О. Г. Нанова, Н. Н. Спасская
К изучению морфологического разнообразия размерных признаков черепа млекопитающих. 1. Соотношение разных форм групповой изменчивости
Стр. 344–354
Резюме. Abstract
Синопсис: Новые методы изучения групповой изменчивости помогают понять природу биологического разнообразия
И. О. Тетевина, В. Г. Онипченко
Долговременная реакция видов растений четырех альпийских сообществ северо-западного Кавказа на перенесение их в новые экологические условия
Стр. 195–206
Резюме. Abstract
Синопсис: Доминанты в новых условиях не выживают
Н. А. Заренков
Опыт приложения неклассических симметрии к природным биоморфам
Стр. 403–423
Резюме. Abstract
Б. М. Миркин, С. М. Ямалов, Л. Г. Наумова
Синантропные растительные сообщества: модели организации и особенности классификации
Стр. 435–443
Резюме. Abstract
Синопсис: «Что вижу, то пою» в геоботанике
А. Т. Терехин, Т. де Мееус, Ж. -Ф. Геган
О мощности некоторых биноминальных модификаций множественного критерия Бонферрони
Стр. 332–340
Резюме. Abstract
И. Я. Павлинов
Филогенетическое мышление в современной биологии
Стр. 19–34
Резюме. Abstract
А. П. Расницын
Классическая и неклассическая систематика: другой взгляд
Стр. 385–388
Резюме. Abstract
Ю. А. Песенко
Есть разница между научной гипотезой и «филогенетической презумпцией» А. П. Расницына (по поводу статьи И. Я. Павлинова, 2005. «0 значении презумпций в филогенетике». Журн. Общ. Биологии. Т. 66. № 5)
Стр. 222–223
Резюме. Abstract
А. П. Расницын
Снова о презумпциях (по поводу дискуссии между И. Я. Павлиновым «О значении презумпций в филогенетике», 2005). Журн. Общ. Биологии. Т. 66 № 5, и Ю. А. Песенко «Есть разница между научной гипотезой и «филогенетической презумпцией» А. П. Расницина», 2006.
Стр. 224–226
Резюме. Abstract
Б. М. Миркин, Л. Г. Наумова
Рецензия на книгу: Крупнейшие ветланды мира. Экология и охрана / под ред. Фрейзера Л. Х. Кедди П А. Кэмбридж, 2005. 488 с.
Стр. 236–239
И. Я. Павлинов
Классическая и неклассическая систематика: где проходит граница?
Стр. 83–106
Резюме. Abstract
М. В. Козлов, С. X. Хёлберт
Мнимые повторности, бесплодные дискуссии и интернациональная сущность науки: ответ Д. В. Татарникову
Стр. 145–152
Резюме. Abstract
Б. М. Миркин
Рецензия на книгу Кафедра геоботаники Московского университета: 75 лет со дня основания
Стр. 75–78
В. В. Акатов, С. Г. Чефранов, Т. В. Акатова
Роль исторических процессов в определении современного видового богатства древесного яруса лесов Западного Кавказа
Стр. 459–470
Резюме. Abstract
Б. М. Миркин
Проблема соотношения непрерывности и дискретности и современная экология
Стр. 522–526
Резюме. Abstract
Б. М. Миркин, Л. Г. Наумова, В. Б. Мартыненко
Рецензия на книгу Восточноевропейские леса: история в голоцене и современность: в 2 кн.
Стр. 363–368
Резюме.
С. Н. Кирпотин
Жизненные формы организмов как паттерны организации и пространственные экологические факторы
Стр. 239–250
Резюме. Abstract
Д. О. Логофет, Е. А. Денисенко, Л. Л. Голубятников
Сукцессии в лесостепи в условиях изменения климата: модельный подход
Стр. 136–145
Резюме. Abstract
Ю. А. Песенко
"Филогенетические презумпции" - могут ли понятия юриспруденции помочь сравнительной биологии?
Стр. 146–163
Резюме. Abstract
Д. В. Леонтьев, А. Ю. Акулов
Экоморфема органического мира: опыт построения
Стр. 500–526
Резюме. Abstract
Д. Б. Гелашвили, Е. В. Чупрунов, Д. И. Иудин
Структурные и биоиндикационные аспекты флуктуирующей асимметрии билатерально-симметричных организмов
Стр. 433–441
Резюме. Abstract
А. И. Кафанов, Е. Э. Борисовец, И. В. Волвенко
О применении кластерного анализа в биогеографических классификациях
Стр. 250–265
Резюме. Abstract
Б. М. Миркин, В. Б. Мартыненко, Л. Г. Наумова
Значение классификации растительности для современной экологии
Стр. 167–177
Резюме. Abstract
Том 77 № 4, Июль-август, 2016
Экспериментально установлено, что длительная адаптация мух Drosophila melanogaster к жизни на корме с повышенным содержанием поваренной соли не сопровождается снижением их приспособленности при развитии на обычном корме. За 20 поколений мухи успешно адаптировались к жизни на «стрессовом» субстрате с NaCl, на котором приспособленность контрольных мух сильно снижена. При этом приспособившиеся к жизни на новом корме мухи по числу выплаживающихся потомков и темпам развития превосходили контрольных также и на обычном корме. Из этого следует, что отбор на устойчивость к развитию на корме с высоким содержанием соли привел к расширению нормы реакции у мух, адаптировавшихся к новым, экстремальным условиям существования. Эта тенденция препятствует разделению исходной популяции в ходе освоения ею новых условий обитания и тормозит процесс дивергенции.
Том 77 № 4, Июль-август, 2016
Исследования следовой активности в зимний период помогают установить пространственные и временные закономерности динамики популяций. Перемещения могут быть вызваны различными классами факторов: расселением, взаимодействием видов, стохастическими причинами. Авторы статьи отслеживали следовую активность лесной куницы, зайца-беляка и обыкновенной белки в Центрально-лесном заповеднике. Удалось показать, что основным параметром, обуславливающим синхронизацию активности хищника и жертвы, является фактор неблагоприятных погодных условий. Пространственно-временная динамика взаимодействующих популяций зависит также от их численности и характеристик местообитания (наличие укрытий).
Том 77 № 4, Июль-август, 2016
Биологический смысл реакции на стресс – мобилизация внутренних ресурсов организма для защиты от угроз, которые неизбежно возникают при взаимодействии животных с внешней средой – холода, голода, нападения хищников. Когда такие воздействия становятся слишком частыми, свободные ресурсы истощаются, и организм оказывается бессилен перед новыми экологическими «вызовами». Возможно, именно это и определяет, в конечном счете, распространение видов. Если это так, то физиологические характеристики, отражающие реакцию организма на стрессирующие воздействия внешней среды, должны различаться у животных, обитающих в благоприятных и неблагоприятных для вида условиях. Чтобы проверить это предположение авторы сравнили реакцию на острое охлаждение у таежного грызуна – красной полевки из популяций, населяющих оптимальные и неоптимальные для вида местообитания юга Западной Сибири. Оказалось, что зверьки, пойманные в оптимальном местообитании – Алтайской горной тайге, демонстрировали более высокую гормональную (неспецифический компонент реагирования, срабатывающий не зависимо от источника стресса), но более низкую метаболическую (непосредственно отвечающую за выработку тепла) реакцию на модельный стрессор, чем полевки из лесостепной популяции окрестностей г. Новосибирска.
МАТЕРИАЛЫ ПО ТЕМАМПоверхность торфа шероховатая от хорошо различимых остатков растений, вода выжимается, как из губки, светлая. При сжатии торф пружинит, не продавливается между пальцами, не грязнит руку
Слабо разложившийся торф
Остатки растений заметны хуже, вода выжимается в большом количестве, мутновато-желтого цвета. При сжатии торф пружинит, слабо продавливается между пальцами, не грязнит руку
Средне и хорошо разложившийся торф
Слегка заметны немногочисленные остатки растений. Масса торфа почти не продавливается в руке, вода отжимается мутными каплями, коричневого цвета. При сжатии торф не пружинит (пластичен), продавливается на 1/3 между пальцами, слабо грязнит руку
Сильно разложившийся торф
Растительные остатки почти неразличимы (преобладает темный гумус). Масса торфа легко продавливается между пальцами, пачкая руку, иногда в виде грязеподобной массы, вода почти не отжимается, темно-коричневого цвета
Alopecurus pratensis Festuca pratensis Elytrigia repens Poa pratensis Agrostis gigantea Carex praecox Poa palustris Carex caespitosa Eleocharis palustris Sanguisorba officinalis Veronica longifolia
Ranunculus acris Ranunculus olyanthemos Symphytum officinale Glechoma hederacea Viola montana Leontodon autumnalis Phleum pratense Iris sibirica Eryngium planum Equisetum pratense Thalictrum flavum
Allium angulosum Rorippa brachicarpa Inula britannica Filipendula ulmaria Peucedanum palustre Rumex acetosa Lysimachia nummularia Leucanthemum vulgare Inula hirta Serratula coronata Galium rubioides Rumex confertus Vicia cracca Lysimachia vulgaris Cirsium setosum Tripolium vulgare Ranunculus auricomus
Calamagrostis epigeios Lathyrus pratensis Mentha arvensis Taraxacum officinale Potentilla anserina Galium album Veronica scutellata Poa angustifolia Rosa majalis Viola canina Hieracium umbellatum Plantago major Prunella vulgaris Sedum telephium Cenolophium fischeri Galium septentrionale Phalaroides arundinacea
Пример описания лесного фитоценоза
№ описания. 1. Автор описания. Соломещ А.И.
Дата описания. 24.06.90.
Местоположение описываемого участка растительности. Башкортостан, Краснокамский район, 1,5 км к В от деревни Николо-Березовка.
Местообитание. выровненный участок
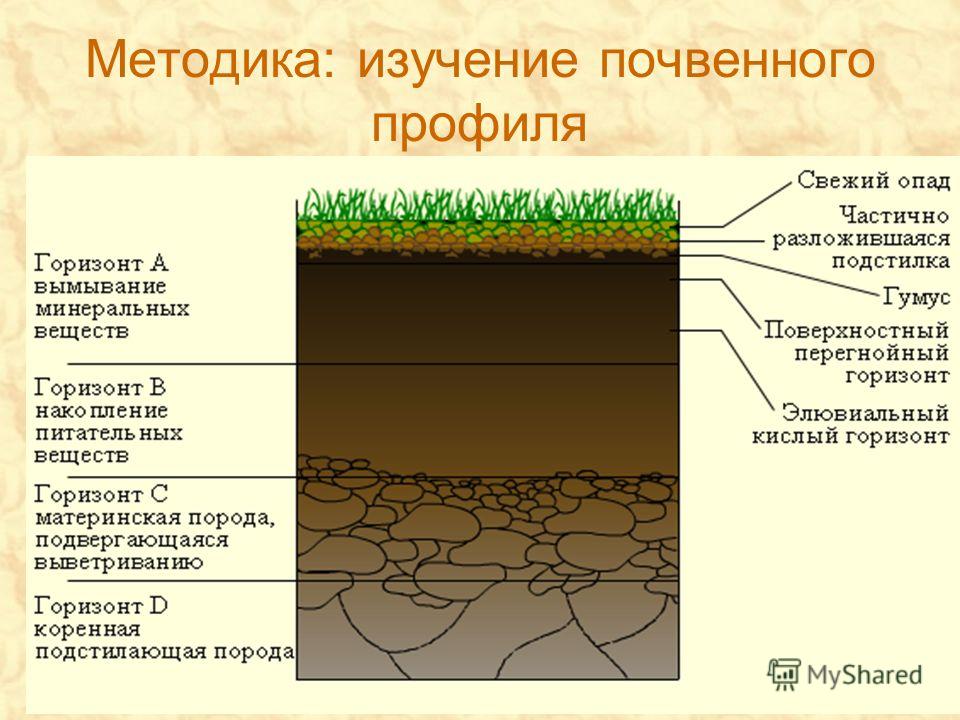
Почва. темно-серая лесная.
Площадь описания. 625 м .
Средняя высота 35 м, максимальная – 40 м.
Средний диаметр ствола 40 см, максимальный – 65 см.
Расстояние между стволами 5 м.
Проективное покрытие 60% (или полнота древостоя 0,6).
Формула древостоя 6С4Е.
Проективное покрытие 10-15%.
Средняя высота 1,5-2 м.
Проективное покрытие 65%.
Средняя высота 20-30 см, максимальная - 70 см.
Проективное покрытие 1-2%.
Число видов общее 66.
Число древесных, кустарниковых и травяных видов 56.
Число видов мхов 10.
Видовой состав описывался на исследованном участке по определителям растительной флоры.
Морфологические особенности деревьев и кустарников на двух участках урочища Чёрный лес (Воронов, 1973)Количество экземпляров растений подсчитывали на двух пробных площадках. Были заложены площадки 25 х 25 м 2. общей площадью каждая 625 м 2. Учитывали те растения которые располагались в пределах площадки. Определяли диаметр растений на высоте груди, по следующим формулам:

P - длина окружности дерева;
Высоту растения определяли глазомерно. Для этого на стволе намечали какую либо точку, находящуюся на высоте 1-3 м, затем, отойдя от дерева, так чтобы был полностью виден весь ствол дерева, мысленно откладывали это расстояние вверх по стволу. Так поступали до тех пор, пока не оставалось некоторое расстояние до вершины. После этого подсчитывали количество отрезков и определяли общую высоту дерева. Полноту древостоя определяли визуально по сомкнутости крон. Если полог леса плотный, и просветы между кронами занимали не более 10 % площади древостоя, такую полноту принимали за 1,0, а последующие ступени уменьшения полноты 1/10 - за 0,9; 0,8; 0,7 и т.д. Сомкнутыми принято называть насаждения с полнотой 0,8-1,0, средней полноты - 0,6 -0,7, редкими - 0,4 - 0,5. Насаждения имеющие полноту менее 0,4, называют рединами.
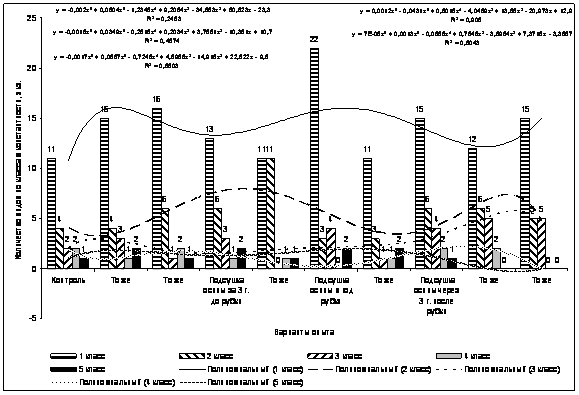
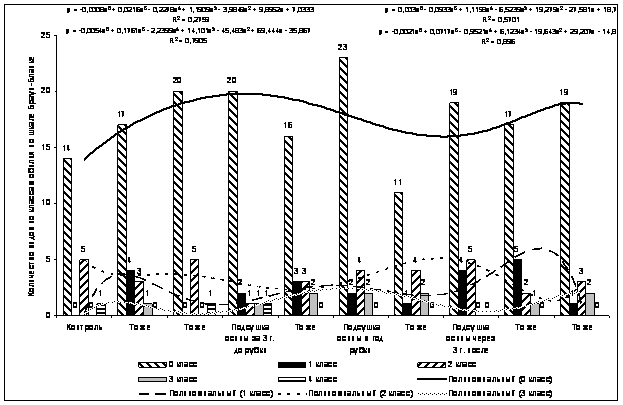
Проведен дисперсионный анализ по методике полевого опыта (с основами статистической обработки результатов исследований) Доспехова Б.А. 1985 г.
Основными статистическими характеристиками количественной изменчивости являлись: средняя арифметическая (Х), дисперсия (S 2 ), стандартное отклонение (S), ошибка выборки (Sх), коэффициент вариации (V) и относительная ошибка выборочной средней (Sх %). В таблицах приведены Х, V, Sх %.
Принципы учета обилия вида (Воронов, 1973)Обилие вида в сообществе - его количество, которое может быть выражено различными показателями: числом особей на единицу площади; массой органического вещества, производимой видом; пространством занимаемым особям вида.
Принцип учета обилия видов определялся глазомерным способом по предложенной шкале Друде (Воронов, 1973).
Были заложены прямоугольники, вытянутые поперек основного направления однородных участков фитоценоза (25х25 м 2 ).
По шкале Друде учитывалось обилие вида:
Soc (soceales) - широко распространено, ему соответствует 4 балла;
Cop (copeaseae) - обильно, соответствует 3 балла;
Sp (изредко рассеянные), ему соответствует 2 балла;
Sol (единично, редко), соответствует 1 балл.
Методика описания ярусности (Воронов, 1973)Ярусностью фитоценоза называют размещение органов растений различных видов на разных высотах над поверхностью почвы. В ходе нашей работы мы выделили пять ярусов и отобразили в гистограмме.
К первому ярусу отнесли высокорослые деревья, ко второму - среднерослые деревья, третий ярус - кустарники, четвертый - травы, в пятый ярус входили мхи и лишайники.
Фенологические наблюдения за лесной растительностью флоры(Воронов, 1973)Мозаичность фитоценоза - это проявление особенностей структуры в пространстве и смена фенологических фаз и аспектов как проявление структуры во времени.
В.В.Алёхин (1961) предложил систему значков, которая и была использована в нашей работе.
- - растение только вигитирует, находится в стадии розетки, начинает давать стебель;
Л - растение выбросило стебель или стрелку и имеет бутоны;
) - растение в фазе расцветания, появляются первые цветки;
О - растение в полном цвету;
С - растение в фазе отцветания;
+ - растение отцвело, но семена еще не созрели и не высыпаются;
# - семена (плоды) созрели и высыпаются (отпадают);
- растение вегетирует после цветения.
Методика описания и классификация растительности методом Браун - Бланке (Миркин, 2001)Последовательность действий при выполнении геоботанического описания следующая. Выбор места описания. Пробные площадки для геоботанических описаний закладывались в однородных (гомогенных) участках растительности, представляющих ранее намеченные в ходе рекогносцировки совокупности.
Выбор размера и формы описываемого участка растительности. Использовались пробные площадки квадратной формы, размером 25х25 м.
Составление описания. Геоботаническое описание проводилось на специальных бланках. Каждое описание содержало указание даты, автора, местоположения (топографической привязки) и местообитания. Для древесной растительности отдельно описывались ее ярусы и их подразделения. Указывалось проективное покрытие, оно не может превышать 100 %.
В описании использовалась шкала обилия Браун-Бланке (Миркин, 2001):
r - вид чрезвычайно редок, покрытие незначительное;
+ - вид редок и имеет малое проективное покрытие;
1 - особей вида много, но покрытие невелико или особи разрежены, но покрытие большое;
2 - число особей вида велико, проективное покрытие 5 - 25 %;
3 - число особей вида любое, проективное покрытие 25 - 50 %;
4 - число особей вида любое проективное покрытие 50 - 75 % ;
5 - число особей вида любое проективное покрытие более 75 %.
Анализ состава флор (Ипатов, 1983)Для анализа флор составляются определенные спектры. В нашей работе проанализировано распределение видов по десяти ведущим семействам по Толмачёву (1986).
Были использованы такие показатели как среднее число видов в роде, среднее число родов в семействах, которые могут получить эволюционную интерплитацию (чем больше родов в семействах, тем они древнее; чем больше видов в родах, тем напротив, они отражают более поздние этапы эволюции.
Если Вы заметили ошибку в тексте выделите слово и нажмите Shift + Enter







